Artigo
Abelha Iratim (Lestrimelitta limao Smith: Apidae, Meliponinae), realmente é danosa às populações de abelhas? Necessita ser eliminada?
Weyder Cristiano Santana1, Geusa Simone de Freitas2 Ivan Paulo Akatsu3, e Ademilson Espencer Egea Soares41, 2, 3 - Departamento de Biologia, FFCLRP-USP, Avenida Bandeirantes, 3900, CEP 14049-901 Ribeirão Preto, SP, Brasil. (wcs@usp.br 1, varzea@usp.br 2, akatsu@usp.br 3)
4 - Departamento de Genética, FMRP-USP, Av. Bandeirantes 3900, CEP 14049-900, Ribeirão Preto, SP, Brasil. (aesoares@fmrp.usp.br) (16) 6023155
Resumo:
Dentre as abelhas indígenas altamente socais, Meliponinae, Lestrimelitta limao (iratim), apresenta distribuição exclusivamente pantropical e se caracteriza por ser cleptoparasita, possui o hábito de saquear o alimento (néctar e pólen) de outras colônias de abelhas. O presente trabalho teve como objetivo verificar a estratégia de ataque de L. limao (abelha limão) a ninhos de outras abelhas sem ferrão, localizadas em cavidades naturais e não naturais (cavidades em construções e em colméias racionais) no Campus da USP de Ribeirão Preto, SP, Brasil. O ataque desta abelha se inicia com a construção de uma lamela de cerume, liberação de feromônios e o comportamento agressivo que evita o pouso e a entrada de abelhas da colônia atacada. As operárias de abelhas limão se mantém no interior da lamela dando continuidade ao ataque e realizando a regulação da temperatura interna durante a noite. A abertura desta lamela pela manhã se dá com a chegada de outras operárias de abelha limão da colônia agressora. Foram observados 5 ataques em colméias racionais de Tetragonisca angustula (jataí) e Frieseomelitta varia (marmelada) e 18 ataques em ninhos localizados em cavidades naturais de diferentes espécies. Verificamos que nestes ataques poucas colméias foram dizimadas pelas abelhas limão ao saquear todo o alimento e cerume (16%). Em condições naturais é possível a convivência entre L. limao (iratim) e outras espécies de abelhas sem que seja necessário destruí-la, ocorrendo um equilíbrio destas populações.
Introdução:
As abelhas eusociais da subfamília Meliponinae, apresentam distribuição pantropical com a maior diversidade de espécies ocorrendo nos neotrópicos e apresentam número estimado acima de 300 espécies (Camargo et al, 1992). À exceção da maior parte das espécies dessas abelhas que buscam seus alimentos nas flores, Lestrimelitta spp (várias espécies) não possui o hábito de coletar pólen e néctar em plantas, sendo cleptobióticas, ou seja, suas operárias pilham ninhos de várias espécies de abelhas da tribo Trigonini e menos comumente Meliponini e Apini (Michener, 1974), sendo que o mel e o pólen pastoso de iratim são tóxicos para o consumo humano (Nogueira-Neto, 1997).
O sucesso no ataque a outras colônias dá-se por liberação de terpenóides voláteis das secreções cefálicas (das glãndulas mandibulares) que provocam a dispersão dos indivíduos da colônia hospedeira e a consequente pilhagem (Francke et al, 2000), por isto o cheiro semelhante a limão que estas abelhas possuem e o nome popular de Abelha limão. As operárias de iratim transportam o mel e o pólen no papo, enquanto o cerume, utilizado na construção do ninho e favos, na tíbia do terceiro par de pernas (Michener, 1946). L. limao (iratim) tem provavelmente sobrevivido formando colônias estáveis capazes de emitir ataques de tamanho efetivo, sendo sua densidade de ninhos baixa (Sakagami et al, 1993).
A entrada do ninho apresenta protuberãncias de cerume que são abertas pelas operárias no período da manhã e fechadas ao anoitecer (Sakagami e Laroca, 1963) (figura 2).
Foi proposto neste estudo verificar a inter-relação de L. limao e outras abelhas, a preferência por espécies, o raio de ataque, a frequência de ocorrência dos ataques e os tipos de ataque.
Material e Métodos:
A área do Campus foi reflorestada a partir de 1946 (Kerr, informação pessoal) onde foram introduzidas algumas espécies de Meliponinae (abelhas indígenas), incluindo L. limao (iratim) em uma área próxima a este em 1991, constituindo-se assim um modelo interessante de estudos da inter-relação e equilíbrio entre as espécies de abelhas.
O estudo dos hábitos cleptobióticos de L limao foi realizado a partir do levantamento dos ninhos de abelhas da subfamília Meliponinae que ocorrem na área de 574,638 hectares do Campus de Ribeirão Preto, São Paulo, Brasil, localizado a 21o05· S e 47o55·W e com altitude máxima de 653 metros (figura 1) (Freitas, 2001). A partir destes dados e com a localização dos ninhos das abelhas iratim, os ninhos de outras espécies de meliponíneos foram acompanhados no sentido de verificar a existência de vestígios de ataques, que são pequenas porções de cerume que aparecem nas entradas dos ninhos das abelhas atacadas (Laroca e Orth, 1984; Bego et al, 1991; Sakagami et al., 1993) e o estado da colônia parasitada ao final do ataque.

Figura 1: Vista aérea do Campus da USP de Ribeirão Preto,
São Paulo.
A área foi percorrida para a localização e posicionamento dos ninhos, utilizando o programa Autocad R14, que permitiu calcular as distãncias interespecíficas e intraespecíficas dos ninhos (Freitas, 2001).
O estudo foi realizado com base em observações diretas incidentais de alguns ataques pela presença da lamela na entrada de várias espécies; e sistematizada de L. limao (iratim) em ninhos de Frieseomelitta varia (marmelada).
Observou-se todo o período do ataque de abelhas limão ao ninho de F. varia instalado em caixa racional. A temperatura no interior da lamela de cerume construída pelas L. limao e a temperatura externa foram medidas a cada 15 minutos durante o período de observação direta.
Resultados e discussão:
Os ninhos de meliponíneos encontram-se na sua maioria localizados em muros das construções, cavidades em troncos de árvores e alguns poucos em caixas racionais de criação (Freitas, 2001).
Os ataques de L. limao foram verificados em ninhos de Tetragonisca angustula (jataí), Nannotrigona testaceicornis (iraí), Plebeia droryana (mirim) e Friesella schrottkyi (mirim preguiça) em colônias sob condições naturais (Freitas, 2001) e T. angustula (jataí) e Frieseomelitta varia (marmelada) em caixas racionais.
Nas colméias foram observadas a presença da lamela construída pelas operárias de L. limao em volta da entrada dos ninhos das espécies atacadas (Bego et al, 1991) (figura 3). Tais lamelas têm por finalidade impedir a entrada de operárias das colônias atacadas e permitir o estabelecimento das abelhas de limão durante a noite, resguardando o ataque, permitindo que este perdure durante a noite e a manutenção da concentração dos feromônios envolvidos na pilhagem.
Os compostos voláteis (terpenóides) secretados pelas glãndulas cefálicas das abelhas limão possuem efeito sobre a comunicação das operárias agredidas (Francke et al, 2000), confundindo-as, desorganizando-as e causando a dispersão destas, permitindo o sucesso do ataque, mesmo com um número baixo de indivíduos agressores observados no início do ataque.

Figura 2: Colônia mãe de L. limao (iratim) localizada
a 200 metros de distãncia da colônia saqueada de
Frieseomelitta varia (marmelada).
Sistematização do ataque:
O início do ataque de L limao (iratim) a F. varia (marmelada) ocorreu às 15:30 do dia 01 de julho de 2003, quando um grupo de 40 a 50 operárias de abelha limão dominou a entrada da colônia de marmelada e iniciou a construção da lamela evitando que qualquer indivíduo dessa colônia pousasse sobre o tubo (figura 3a, 3b, 3c). Tais abelhas chegaram com cera da colônia agressora.
As operárias de marmelada, numa tentativa de defesa, fecharam a boca de entrada por dentro com resina (figura 3a, 3b, 3c). A lamela de cerume foi finalizada até 18:30 horas do mesmo dia do início do ataque (figura 3d).
Algumas operárias de iratim foram marcadas e permaneceram no interior da lamela durante a noite (figura 3d, 3f, 3k).
A abertura da lamela na manhã do dia seguinte (02/07/2003) ocorreu as 8:45 horas, com a saída da primeira abelha às 8:53 horas em direção a colônia de L. limao (figura 3h). E apesar de pequena, a abertura na lamela possui as mesmas características da boca de entrada da colméia de abelhas limão, com operárias em guarda (figura 2, 3e, 3k). A partir deste horário iniciou a chegada de operárias de abelhas limão em um pequeno número, não mais que 2 a 3 indivíduos a cada 15 minutos, aproximadamente.
Operária de abelha limão pousou sobre a lamela e depositou uma gotícula de substãncia de cheiro para facilitar a orientação de outras operárias que vinham da colônia de Lestrimelitta (figura 3f, 3g). Da mesma forma, outras operárias de iratim depositaram resina sobre o cerume, marcando o local visualmente (figura 3g, 3h, 3k).
Um grande número de operárias de L. limao vindos da colméia foi observado às 11:47 horas, o que caracterizou o início de um ataque em massa (figura 3i). Sendo o período de maior atividade de vôo das operárias de iratim, durante o saque, entre 11:00 e 16:00 horas, que alterna picos de intenso e baixo fluxo de operárias de abelhas limão (31-67 abelhas/minuto saindo e 15-30 chegando) (figura 3i)
A partir deste ponto, também se intensificou a remoção de operárias mortas de F. varia por operárias de limão que as jogaram pela entrada da lamela ao chão.
Na colônia de Lestrimelitta foi observado a chegada de operárias marcadas na colméia de F. varia com tinta, comprovando a origem destas. Todas as operárias campeiras que retornavam estavam com o abdômen distendido, voando baixo, que caracterizou estarem pesadas; além disso, carregavam cerume na corbícula rudimentar, confirmando o saque na colônia de marmelada. Algumas operárias de L. limao que saiam foram coletadas e o conteúdo do papo foi examinado, sendo verificado que estavam carregando mel, e/ou uma mistura de mel e pólen no papo, e também cerume aderido às corbículas rudimentares. Tais recursos, principalmente o cerume parece ser muito importante para a ampliação das colônias de L. limao.
Em torno de 15 operárias de abelha limão, sempre permaneciam trabalhando na entrada da lamela e ao mesmo tempo guardando a boca do ninho (figura 3e e 3k).
Durante o período de saque à colônia de marmelada surgiram formigas, forídeos (diptero parasita de ninhos de abehas indígenas) e algumas abelhas: Apis mellifera (africanizada), Plebeia droryana (mirim), Oxytrigona tataíra (caga-fogo), Scaptotrigona depilis (tubiba) e Trigona spinipes (abelha-cachorro ou arapuá). Estas pousavam próximo à lamela atraídas pelo alvoroço, e/ou algum cheiro de alimento da colônia, e/ou pelo néctar depositado pelas operárias campeiras de marmelada sobre a colméia (figura 3k).
A boca da lamela da abelha limão foi fechada entre 17:00 e 18:30 horas (figura 3d), mesmo período verificado na colônia de abelha limão (figura 2).
A partir do dia 04/07/2003, as operárias de limão estavam transportando da colônia atacada mais cerume na corbícula rudimentar que mel e pólen no papo, observados nos primeiros dias.
No dia 05/07/2003, algumas operárias de marmelada (aproximadamente 20 indivíduos) ficaram sobre o tubo, antes deste ser aberto pelas abelhas limão, defendendo-o do pouso de algumas operárias de iratim. Acredita-se que tais operárias campeiras de marmelada que estavam fora da colméia no início da pilhagem, não puderam retornar a colméia e ficaram para fora, provavelmente pernoitaram nas flores e voltaram, pois estavam marcadas, durante todo este período de observação, bem como traziam néctar que depositavam sobre a lamela ou sobre a caixa. Após a abertura da lamela, iniciava-se um comportamento de vôo em zigue-zague pelas operárias de L. limao ao verificarem que algumas operárias de F. varia estavam tentado entrar no tubo.
No quinto dia (06/07/2003), entre 13:55 à 14:30 horas, chegaram mais operárias de abelha limão; e a partir de 14:35 horas saíram mais operárias que chegaram, até 16:55 horas, indicando a finalização do saque. A partir das 17:00 horas as operárias marcadas de F. varia, que voavam próximo à colméia, passaram a entrar na lamela e não encontrando operárias de L. limao em seu interior, passaram a destruir o invólucro deixado pela abelha limão (figura 3l).
Ao abrir a colméia de marmelada, observou-se que a abelha limão carregou todo o alimento (pólen e mel) dos potes e o cerume das pilastras e das células novas de cria, bem como o alimento nele contido (figura 3n). A rainha fisogástrica foi observada com algumas abelhas jovens e células de crias mais velhas intactas.
As operárias de F. varia construíram um tubo na parte interna da caixa racional na tentativa de impedir a entrada da L. limao em uma estratégia de defesa (figura 3m).
A temperatura média no interior da lamela foi maior que a temperatura do ambiente em 9,225 °C durante o período da manhã (com sol), 2,125 °C no período da tarde (sem sol). Durante a noite, a temperatura no interior da lamela estava 2,375 °C maior que o exterior, caracterizando uma regulação térmica devido a atividade das abelhas em seu interior (figura 4).
A lamela construída pelas abelhas limão mudou gradativamente de coloração com as constantes marcações destas abelhas, bem como com a regurgitação de néctar pelas operárias de marmelada (figura 3d, 3e, 3f, 3j, 3k).
Esta colônia de F. varia sobreviveu, recuperou-se e foi novamente atacada pela colônia de L. limao no dia 08/08/2003 e no dia 20/08/2003. Neste dois saques posteriores, sobreviveram e se recuperaram.

h: Saída de algumas operárias de Iratim em direção a colônia de origem (seta). i: Grande número de operárias de L. limao
chegando para o saque; a seta indica a sonda de temperatura exterior. j: Início da abertura da boca da lamela. k: Operárias
de L. limao marcadas com tinta da cor laranja; boca da lamela com guardas, sendo marcada (seta) e aumentada; lamela
mais escurecida. l: Destruição da lamela pelas operárias de F. varia após o saque. m: Tubo de cerume construído pelas
operárias de F. varia no interior da caixa para se proteger do ataque de L. limao. n: Aspecto do interior da colméia de
F. varia após o saque de todo alimento, cerume e do conteúdo das células de crias novas (seta).
Ataques incidentais:
Outro ataque de L. limao em F. varia foi observado 5 dias após esta pilhagem, em outra colônia muito forte de marmelada (muitos indivíduos, crias e alimentos) instalada em caixa racional, sendo que este não obteve sucesso, devido à intervenção (abertura da caixa) para saber qual o estado geral desta colônia; contudo a lamela estava quase finalizada, indicando que L. limao consegue iniciar um ataque com poucos indivíduos, estabelecendo dominãncia pela ação dos feromônios por ela produzidos. Foi observado que existiam alguns indivíduos marcados, sendo portanto, da mesma colônia do ataque anteriormente descrito.
Foram localizados 5 ninhos de L. limao, estando 3 em cavidades de paredes de alvenaria e 2 em troncos de árvores, com distãncia de 396 metros entre eles e em uma altura máxima de 2,30 metros. Com densidade ninhos baixa, L. limao está em equilíbrio com a população das outras abelhas, tendo em vista a não explosão populacional destas a partir da sua introdução em 1991 em área próxima.
A maior frequência de ataques (70%) foi a Tetragonisca angustula (jataí), contudo esta espécie e N. testaceicornis (iraí) sofreram vários ataques e após estes foram dizimadas (16%). Assim, nem todos os ataques de L. limao resultaram na extinção das colônias, semelhante ao verificado na literatura (Bego et al, 1991), sendo iraí a que mais sofreu com os ataques. Neste trabalho, os ninhos de iraí constituíram o local para o estabelecimento (reocupação) da nova colméia de L. limao. Tal fato caracteriza a suscetibilidade desta abelha ao ataque e indica que a estrutura do ninho é mais atrativa à abelha limão, podendo ser um indicio do processo de enxameagem de iratim.
As espécies com maior densidade de ninhos não foram as mais atacadas e sim algumas espécies preferenciais, corroborando os trabalhos de Bego et al (1991) e Sakagami et al (1993), onde N. testaceicornis (irai) (Michener,1946) e Scaptotrigona depilis (tubiba) sofreram o maior número de ataques (Sakagami et al, 1993) e T. angustula (jataí) foi a menos preferida (Bego et al, 1991), estando relacionado ao comportamento de defesa de cada espécie e de jataí em especial (Wittmann, 1985 e Wittmann et al., 1990).
O ataque de saque foi realizado somente na colônia de F. varia, não sendo observado no raio de 200m o ataque desta colônia de L. limao a outras espécies de abelhas, comumente relatado na literatura (Michener, 1974; Laroca e Orth, 1984; Bego et al, 1991), podendo indicar quão populosa seria uma colônia de abelha limão. Contudo, um ataque de saque posterior em outra colméia de F. varia e outros ataques sucessivos na primeira colônia, caracterizaram a freqüência dos ataques de abelha limão ou a necessidade de alimento por estas durante o período de Inverno no mês de Julho.
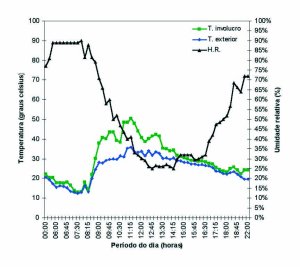
Figura 4: Gráfico contendo as médias das temperaturas do
interior da lamela (sonda), as médias das temperaturas do exterior
e umidade relativa. Dados coletados durante a observação
do saque de L. limao (iratim) em F. varia (marmelada).
Conclusão:
Um parasito que destrói por completo o hospedeiro tende a desaparecer com o tempo se a taxa de multiplicação (reprodução) do hospedeiro for semelhante ou igual ao do parasito. Ou seja, o que deve ser relacionado é o grau do dano causado ao hospedeiro e a taxa de multiplicação deste em relação ao do parasito, bem como a densidade destas populações. Observou-se neste trabalho, baixo número de ninhos de L. limao, estando em acordo com o verificado por outros autores (Sakagami et al, 1993; Bego et al, 1991). A densidade dos ninhos de L. limao (iratim) está em equilíbrio com a população das outras abelhas, evidenciado pela não explosão populacional destas a partir da sua introdução em 1991. Não necessitando, assim, de serem exterminadas.
Muitos ninhos foram atacados, mas não foram destruídos, indicando que estas abelhas não são prejudiciais como lhes é atribuído. Este argumento tem sido utilizado por meliponicultores que sistematicamente efetuam a extinção dos ninhos encontrados. L. limao (iratim) não destrói o ninho hospedeiro em condições naturais no primeiro ataque, mas quando este está muito fraco, ou seja, poucos indivíduos e cria, comum após o terceiro ou quarto ataque sucessivo, caso estas não se recuperem, as colônias podem ser destruídas. O mesmo pode acontecer com abelhas criadas em caixas racionais, devido à falta de manejo adequado e ao estado geral da colônia não ser o ideal, semelhante a uma colônia em condição natural, pois a mesma está submetida à constante stress (Nogueira-Neto,1999).
Agradecimentos:
Prefeitura Municipal de Ribeirão Preto, pelo empréstimo das ortofotos do Campus da USP de Ribeirão Preto. CAPES pelo apoio financeiro. Faculdade de Filosofia Ciências e Letras de Ribeirão Preto e à Faculdade de Medicina de Ribeirão Preto pelo suporte.
Bibliografia:
Bego L R, Zucchi R, Mateus S. 1991. Notas sobre a estratégia alimentar (cleptobiose) de Lestrimelitta limao Smith (Hymenoptera, Apidae, Meliponinae). Naturalia, 16:119-127.
Camargo J M F, Pedro S R M. 1992. Systematics, phylogeny and biogeography of Meliponinae, a mini review. Apidologie, 23:509-522.
Francke W, Lubke G, Schoder W, Reckziegel A, Imperatriz-Fonseca V, Kleinert A, Engels E, Hartfelder K, Radtke R, Engels W. 2000. Identification of oxygen containing volatiles in cephalic secretions of workers of Brazilian stingless bees. Journal Brazilian Chemistry Society, 11(6):562-571.
Freitas G S. 2001. Levantamento de ninhos de meliponíneos (Hymenoptera, Apidae) em área urbana: Campus da USP, Ribeirão Preto/SP. Dissertação de Mestrado. Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, USP. Ribeirão Preto, SP.
Laroca, S. & A. I. Orth. 1984. Pilhagem de um ninho de Plebeia catamarcensis meridionalis por Lestrimelitta limao (Apidae, Meliponinae) em Itapiranga, SC, Sul do Brasil. Dusenia 14 (3): 123-127.
Michener, C D. 1946. Notes on the habits of some panamanian stingless bees (Hymenoptera, Apidae). Journal of the New York Entomological Society, LIV(3): 179-197.
Michener, C D. 1974. The social behavior of the bees: A comparative study. Harvard University Press. 404 p.
Nogueira-Neto, P. 1997. Vida e criação de abelhas indígenas sem ferrão. Nogueirapis, São Paulo. 446p.
Nogueira-Neto, P. 1999. Stressful situation and their consequences on sex determination and on the behavior of stingless bees (Meliponinae, Apidae, Hymenoptera). Revista de Ecologia, 1 (1): 65-68.
Sakagami S F, Laroca S. 1963. Additional observations on the habits of the cleptobiotic stingless bees the genus Lestrimelitta Friese. Journal of Faculty Science of Hokkaido University, Seri I. 15:319-339.
Sakagami S F, Roubik D W, Zucchi R. 1993. Ethology of the robber stingless bee, Lestrimelitta limao (Hymenoptera: Apidae). Sociobiology, 21(2): 237-277.
Wittmann, D. 1985. Aerial defense of the nest by workers of the stingless bee Trigona (Tetragonisca) angustula (Latreille) (Hymenoptera, Apidae). Behaviour Ecology and Sociobiolgy 16:111-114.
Wittmann, D.; Radtke, R.; Zeil, J.; Lubke, G. and Francke, W. 1990. Robber bees (Lestrimelitta limao) and their host chemical and visual cues in nest defense by Trigona (Tetragonisca) angustula (Apidae: Meliponinae). Journal of Chemical Ecology 16(2):631-641.
Dentre as abelhas indígenas altamente socais, Meliponinae, Lestrimelitta limao (iratim), apresenta distribuição exclusivamente pantropical e se caracteriza por ser cleptoparasita, possui o hábito de saquear o alimento (néctar e pólen) de outras colônias de abelhas. O presente trabalho teve como objetivo verificar a estratégia de ataque de L. limao (abelha limão) a ninhos de outras abelhas sem ferrão, localizadas em cavidades naturais e não naturais (cavidades em construções e em colméias racionais) no Campus da USP de Ribeirão Preto, SP, Brasil. O ataque desta abelha se inicia com a construção de uma lamela de cerume, liberação de feromônios e o comportamento agressivo que evita o pouso e a entrada de abelhas da colônia atacada. As operárias de abelhas limão se mantém no interior da lamela dando continuidade ao ataque e realizando a regulação da temperatura interna durante a noite. A abertura desta lamela pela manhã se dá com a chegada de outras operárias de abelha limão da colônia agressora. Foram observados 5 ataques em colméias racionais de Tetragonisca angustula (jataí) e Frieseomelitta varia (marmelada) e 18 ataques em ninhos localizados em cavidades naturais de diferentes espécies. Verificamos que nestes ataques poucas colméias foram dizimadas pelas abelhas limão ao saquear todo o alimento e cerume (16%). Em condições naturais é possível a convivência entre L. limao (iratim) e outras espécies de abelhas sem que seja necessário destruí-la, ocorrendo um equilíbrio destas populações.
Introdução:
As abelhas eusociais da subfamília Meliponinae, apresentam distribuição pantropical com a maior diversidade de espécies ocorrendo nos neotrópicos e apresentam número estimado acima de 300 espécies (Camargo et al, 1992). À exceção da maior parte das espécies dessas abelhas que buscam seus alimentos nas flores, Lestrimelitta spp (várias espécies) não possui o hábito de coletar pólen e néctar em plantas, sendo cleptobióticas, ou seja, suas operárias pilham ninhos de várias espécies de abelhas da tribo Trigonini e menos comumente Meliponini e Apini (Michener, 1974), sendo que o mel e o pólen pastoso de iratim são tóxicos para o consumo humano (Nogueira-Neto, 1997).
O sucesso no ataque a outras colônias dá-se por liberação de terpenóides voláteis das secreções cefálicas (das glãndulas mandibulares) que provocam a dispersão dos indivíduos da colônia hospedeira e a consequente pilhagem (Francke et al, 2000), por isto o cheiro semelhante a limão que estas abelhas possuem e o nome popular de Abelha limão. As operárias de iratim transportam o mel e o pólen no papo, enquanto o cerume, utilizado na construção do ninho e favos, na tíbia do terceiro par de pernas (Michener, 1946). L. limao (iratim) tem provavelmente sobrevivido formando colônias estáveis capazes de emitir ataques de tamanho efetivo, sendo sua densidade de ninhos baixa (Sakagami et al, 1993).
A entrada do ninho apresenta protuberãncias de cerume que são abertas pelas operárias no período da manhã e fechadas ao anoitecer (Sakagami e Laroca, 1963) (figura 2).
Foi proposto neste estudo verificar a inter-relação de L. limao e outras abelhas, a preferência por espécies, o raio de ataque, a frequência de ocorrência dos ataques e os tipos de ataque.
Material e Métodos:
A área do Campus foi reflorestada a partir de 1946 (Kerr, informação pessoal) onde foram introduzidas algumas espécies de Meliponinae (abelhas indígenas), incluindo L. limao (iratim) em uma área próxima a este em 1991, constituindo-se assim um modelo interessante de estudos da inter-relação e equilíbrio entre as espécies de abelhas.
O estudo dos hábitos cleptobióticos de L limao foi realizado a partir do levantamento dos ninhos de abelhas da subfamília Meliponinae que ocorrem na área de 574,638 hectares do Campus de Ribeirão Preto, São Paulo, Brasil, localizado a 21o05· S e 47o55·W e com altitude máxima de 653 metros (figura 1) (Freitas, 2001). A partir destes dados e com a localização dos ninhos das abelhas iratim, os ninhos de outras espécies de meliponíneos foram acompanhados no sentido de verificar a existência de vestígios de ataques, que são pequenas porções de cerume que aparecem nas entradas dos ninhos das abelhas atacadas (Laroca e Orth, 1984; Bego et al, 1991; Sakagami et al., 1993) e o estado da colônia parasitada ao final do ataque.
Figura 1: Vista aérea do Campus da USP de Ribeirão Preto,
São Paulo.
A área foi percorrida para a localização e posicionamento dos ninhos, utilizando o programa Autocad R14, que permitiu calcular as distãncias interespecíficas e intraespecíficas dos ninhos (Freitas, 2001).
O estudo foi realizado com base em observações diretas incidentais de alguns ataques pela presença da lamela na entrada de várias espécies; e sistematizada de L. limao (iratim) em ninhos de Frieseomelitta varia (marmelada).
Observou-se todo o período do ataque de abelhas limão ao ninho de F. varia instalado em caixa racional. A temperatura no interior da lamela de cerume construída pelas L. limao e a temperatura externa foram medidas a cada 15 minutos durante o período de observação direta.
Resultados e discussão:
Os ninhos de meliponíneos encontram-se na sua maioria localizados em muros das construções, cavidades em troncos de árvores e alguns poucos em caixas racionais de criação (Freitas, 2001).
Os ataques de L. limao foram verificados em ninhos de Tetragonisca angustula (jataí), Nannotrigona testaceicornis (iraí), Plebeia droryana (mirim) e Friesella schrottkyi (mirim preguiça) em colônias sob condições naturais (Freitas, 2001) e T. angustula (jataí) e Frieseomelitta varia (marmelada) em caixas racionais.
Nas colméias foram observadas a presença da lamela construída pelas operárias de L. limao em volta da entrada dos ninhos das espécies atacadas (Bego et al, 1991) (figura 3). Tais lamelas têm por finalidade impedir a entrada de operárias das colônias atacadas e permitir o estabelecimento das abelhas de limão durante a noite, resguardando o ataque, permitindo que este perdure durante a noite e a manutenção da concentração dos feromônios envolvidos na pilhagem.
Os compostos voláteis (terpenóides) secretados pelas glãndulas cefálicas das abelhas limão possuem efeito sobre a comunicação das operárias agredidas (Francke et al, 2000), confundindo-as, desorganizando-as e causando a dispersão destas, permitindo o sucesso do ataque, mesmo com um número baixo de indivíduos agressores observados no início do ataque.
Figura 2: Colônia mãe de L. limao (iratim) localizada
a 200 metros de distãncia da colônia saqueada de
Frieseomelitta varia (marmelada).
O início do ataque de L limao (iratim) a F. varia (marmelada) ocorreu às 15:30 do dia 01 de julho de 2003, quando um grupo de 40 a 50 operárias de abelha limão dominou a entrada da colônia de marmelada e iniciou a construção da lamela evitando que qualquer indivíduo dessa colônia pousasse sobre o tubo (figura 3a, 3b, 3c). Tais abelhas chegaram com cera da colônia agressora.
As operárias de marmelada, numa tentativa de defesa, fecharam a boca de entrada por dentro com resina (figura 3a, 3b, 3c). A lamela de cerume foi finalizada até 18:30 horas do mesmo dia do início do ataque (figura 3d).
Algumas operárias de iratim foram marcadas e permaneceram no interior da lamela durante a noite (figura 3d, 3f, 3k).
A abertura da lamela na manhã do dia seguinte (02/07/2003) ocorreu as 8:45 horas, com a saída da primeira abelha às 8:53 horas em direção a colônia de L. limao (figura 3h). E apesar de pequena, a abertura na lamela possui as mesmas características da boca de entrada da colméia de abelhas limão, com operárias em guarda (figura 2, 3e, 3k). A partir deste horário iniciou a chegada de operárias de abelhas limão em um pequeno número, não mais que 2 a 3 indivíduos a cada 15 minutos, aproximadamente.
Operária de abelha limão pousou sobre a lamela e depositou uma gotícula de substãncia de cheiro para facilitar a orientação de outras operárias que vinham da colônia de Lestrimelitta (figura 3f, 3g). Da mesma forma, outras operárias de iratim depositaram resina sobre o cerume, marcando o local visualmente (figura 3g, 3h, 3k).
Um grande número de operárias de L. limao vindos da colméia foi observado às 11:47 horas, o que caracterizou o início de um ataque em massa (figura 3i). Sendo o período de maior atividade de vôo das operárias de iratim, durante o saque, entre 11:00 e 16:00 horas, que alterna picos de intenso e baixo fluxo de operárias de abelhas limão (31-67 abelhas/minuto saindo e 15-30 chegando) (figura 3i)
A partir deste ponto, também se intensificou a remoção de operárias mortas de F. varia por operárias de limão que as jogaram pela entrada da lamela ao chão.
Na colônia de Lestrimelitta foi observado a chegada de operárias marcadas na colméia de F. varia com tinta, comprovando a origem destas. Todas as operárias campeiras que retornavam estavam com o abdômen distendido, voando baixo, que caracterizou estarem pesadas; além disso, carregavam cerume na corbícula rudimentar, confirmando o saque na colônia de marmelada. Algumas operárias de L. limao que saiam foram coletadas e o conteúdo do papo foi examinado, sendo verificado que estavam carregando mel, e/ou uma mistura de mel e pólen no papo, e também cerume aderido às corbículas rudimentares. Tais recursos, principalmente o cerume parece ser muito importante para a ampliação das colônias de L. limao.
Em torno de 15 operárias de abelha limão, sempre permaneciam trabalhando na entrada da lamela e ao mesmo tempo guardando a boca do ninho (figura 3e e 3k).
Durante o período de saque à colônia de marmelada surgiram formigas, forídeos (diptero parasita de ninhos de abehas indígenas) e algumas abelhas: Apis mellifera (africanizada), Plebeia droryana (mirim), Oxytrigona tataíra (caga-fogo), Scaptotrigona depilis (tubiba) e Trigona spinipes (abelha-cachorro ou arapuá). Estas pousavam próximo à lamela atraídas pelo alvoroço, e/ou algum cheiro de alimento da colônia, e/ou pelo néctar depositado pelas operárias campeiras de marmelada sobre a colméia (figura 3k).
A boca da lamela da abelha limão foi fechada entre 17:00 e 18:30 horas (figura 3d), mesmo período verificado na colônia de abelha limão (figura 2).
A partir do dia 04/07/2003, as operárias de limão estavam transportando da colônia atacada mais cerume na corbícula rudimentar que mel e pólen no papo, observados nos primeiros dias.
No dia 05/07/2003, algumas operárias de marmelada (aproximadamente 20 indivíduos) ficaram sobre o tubo, antes deste ser aberto pelas abelhas limão, defendendo-o do pouso de algumas operárias de iratim. Acredita-se que tais operárias campeiras de marmelada que estavam fora da colméia no início da pilhagem, não puderam retornar a colméia e ficaram para fora, provavelmente pernoitaram nas flores e voltaram, pois estavam marcadas, durante todo este período de observação, bem como traziam néctar que depositavam sobre a lamela ou sobre a caixa. Após a abertura da lamela, iniciava-se um comportamento de vôo em zigue-zague pelas operárias de L. limao ao verificarem que algumas operárias de F. varia estavam tentado entrar no tubo.
No quinto dia (06/07/2003), entre 13:55 à 14:30 horas, chegaram mais operárias de abelha limão; e a partir de 14:35 horas saíram mais operárias que chegaram, até 16:55 horas, indicando a finalização do saque. A partir das 17:00 horas as operárias marcadas de F. varia, que voavam próximo à colméia, passaram a entrar na lamela e não encontrando operárias de L. limao em seu interior, passaram a destruir o invólucro deixado pela abelha limão (figura 3l).
Ao abrir a colméia de marmelada, observou-se que a abelha limão carregou todo o alimento (pólen e mel) dos potes e o cerume das pilastras e das células novas de cria, bem como o alimento nele contido (figura 3n). A rainha fisogástrica foi observada com algumas abelhas jovens e células de crias mais velhas intactas.
As operárias de F. varia construíram um tubo na parte interna da caixa racional na tentativa de impedir a entrada da L. limao em uma estratégia de defesa (figura 3m).
A temperatura média no interior da lamela foi maior que a temperatura do ambiente em 9,225 °C durante o período da manhã (com sol), 2,125 °C no período da tarde (sem sol). Durante a noite, a temperatura no interior da lamela estava 2,375 °C maior que o exterior, caracterizando uma regulação térmica devido a atividade das abelhas em seu interior (figura 4).
A lamela construída pelas abelhas limão mudou gradativamente de coloração com as constantes marcações destas abelhas, bem como com a regurgitação de néctar pelas operárias de marmelada (figura 3d, 3e, 3f, 3j, 3k).
Esta colônia de F. varia sobreviveu, recuperou-se e foi novamente atacada pela colônia de L. limao no dia 08/08/2003 e no dia 20/08/2003. Neste dois saques posteriores, sobreviveram e se recuperaram.
h: Saída de algumas operárias de Iratim em direção a colônia de origem (seta). i: Grande número de operárias de L. limao
chegando para o saque; a seta indica a sonda de temperatura exterior. j: Início da abertura da boca da lamela. k: Operárias
de L. limao marcadas com tinta da cor laranja; boca da lamela com guardas, sendo marcada (seta) e aumentada; lamela
mais escurecida. l: Destruição da lamela pelas operárias de F. varia após o saque. m: Tubo de cerume construído pelas
operárias de F. varia no interior da caixa para se proteger do ataque de L. limao. n: Aspecto do interior da colméia de
F. varia após o saque de todo alimento, cerume e do conteúdo das células de crias novas (seta).
Outro ataque de L. limao em F. varia foi observado 5 dias após esta pilhagem, em outra colônia muito forte de marmelada (muitos indivíduos, crias e alimentos) instalada em caixa racional, sendo que este não obteve sucesso, devido à intervenção (abertura da caixa) para saber qual o estado geral desta colônia; contudo a lamela estava quase finalizada, indicando que L. limao consegue iniciar um ataque com poucos indivíduos, estabelecendo dominãncia pela ação dos feromônios por ela produzidos. Foi observado que existiam alguns indivíduos marcados, sendo portanto, da mesma colônia do ataque anteriormente descrito.
Foram localizados 5 ninhos de L. limao, estando 3 em cavidades de paredes de alvenaria e 2 em troncos de árvores, com distãncia de 396 metros entre eles e em uma altura máxima de 2,30 metros. Com densidade ninhos baixa, L. limao está em equilíbrio com a população das outras abelhas, tendo em vista a não explosão populacional destas a partir da sua introdução em 1991 em área próxima.
A maior frequência de ataques (70%) foi a Tetragonisca angustula (jataí), contudo esta espécie e N. testaceicornis (iraí) sofreram vários ataques e após estes foram dizimadas (16%). Assim, nem todos os ataques de L. limao resultaram na extinção das colônias, semelhante ao verificado na literatura (Bego et al, 1991), sendo iraí a que mais sofreu com os ataques. Neste trabalho, os ninhos de iraí constituíram o local para o estabelecimento (reocupação) da nova colméia de L. limao. Tal fato caracteriza a suscetibilidade desta abelha ao ataque e indica que a estrutura do ninho é mais atrativa à abelha limão, podendo ser um indicio do processo de enxameagem de iratim.
As espécies com maior densidade de ninhos não foram as mais atacadas e sim algumas espécies preferenciais, corroborando os trabalhos de Bego et al (1991) e Sakagami et al (1993), onde N. testaceicornis (irai) (Michener,1946) e Scaptotrigona depilis (tubiba) sofreram o maior número de ataques (Sakagami et al, 1993) e T. angustula (jataí) foi a menos preferida (Bego et al, 1991), estando relacionado ao comportamento de defesa de cada espécie e de jataí em especial (Wittmann, 1985 e Wittmann et al., 1990).
O ataque de saque foi realizado somente na colônia de F. varia, não sendo observado no raio de 200m o ataque desta colônia de L. limao a outras espécies de abelhas, comumente relatado na literatura (Michener, 1974; Laroca e Orth, 1984; Bego et al, 1991), podendo indicar quão populosa seria uma colônia de abelha limão. Contudo, um ataque de saque posterior em outra colméia de F. varia e outros ataques sucessivos na primeira colônia, caracterizaram a freqüência dos ataques de abelha limão ou a necessidade de alimento por estas durante o período de Inverno no mês de Julho.
Figura 4: Gráfico contendo as médias das temperaturas do
interior da lamela (sonda), as médias das temperaturas do exterior
e umidade relativa. Dados coletados durante a observação
do saque de L. limao (iratim) em F. varia (marmelada).
Um parasito que destrói por completo o hospedeiro tende a desaparecer com o tempo se a taxa de multiplicação (reprodução) do hospedeiro for semelhante ou igual ao do parasito. Ou seja, o que deve ser relacionado é o grau do dano causado ao hospedeiro e a taxa de multiplicação deste em relação ao do parasito, bem como a densidade destas populações. Observou-se neste trabalho, baixo número de ninhos de L. limao, estando em acordo com o verificado por outros autores (Sakagami et al, 1993; Bego et al, 1991). A densidade dos ninhos de L. limao (iratim) está em equilíbrio com a população das outras abelhas, evidenciado pela não explosão populacional destas a partir da sua introdução em 1991. Não necessitando, assim, de serem exterminadas.
Muitos ninhos foram atacados, mas não foram destruídos, indicando que estas abelhas não são prejudiciais como lhes é atribuído. Este argumento tem sido utilizado por meliponicultores que sistematicamente efetuam a extinção dos ninhos encontrados. L. limao (iratim) não destrói o ninho hospedeiro em condições naturais no primeiro ataque, mas quando este está muito fraco, ou seja, poucos indivíduos e cria, comum após o terceiro ou quarto ataque sucessivo, caso estas não se recuperem, as colônias podem ser destruídas. O mesmo pode acontecer com abelhas criadas em caixas racionais, devido à falta de manejo adequado e ao estado geral da colônia não ser o ideal, semelhante a uma colônia em condição natural, pois a mesma está submetida à constante stress (Nogueira-Neto,1999).
Agradecimentos:
Prefeitura Municipal de Ribeirão Preto, pelo empréstimo das ortofotos do Campus da USP de Ribeirão Preto. CAPES pelo apoio financeiro. Faculdade de Filosofia Ciências e Letras de Ribeirão Preto e à Faculdade de Medicina de Ribeirão Preto pelo suporte.
Bibliografia:
Bego L R, Zucchi R, Mateus S. 1991. Notas sobre a estratégia alimentar (cleptobiose) de Lestrimelitta limao Smith (Hymenoptera, Apidae, Meliponinae). Naturalia, 16:119-127.
Camargo J M F, Pedro S R M. 1992. Systematics, phylogeny and biogeography of Meliponinae, a mini review. Apidologie, 23:509-522.
Francke W, Lubke G, Schoder W, Reckziegel A, Imperatriz-Fonseca V, Kleinert A, Engels E, Hartfelder K, Radtke R, Engels W. 2000. Identification of oxygen containing volatiles in cephalic secretions of workers of Brazilian stingless bees. Journal Brazilian Chemistry Society, 11(6):562-571.
Freitas G S. 2001. Levantamento de ninhos de meliponíneos (Hymenoptera, Apidae) em área urbana: Campus da USP, Ribeirão Preto/SP. Dissertação de Mestrado. Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, USP. Ribeirão Preto, SP.
Laroca, S. & A. I. Orth. 1984. Pilhagem de um ninho de Plebeia catamarcensis meridionalis por Lestrimelitta limao (Apidae, Meliponinae) em Itapiranga, SC, Sul do Brasil. Dusenia 14 (3): 123-127.
Michener, C D. 1946. Notes on the habits of some panamanian stingless bees (Hymenoptera, Apidae). Journal of the New York Entomological Society, LIV(3): 179-197.
Michener, C D. 1974. The social behavior of the bees: A comparative study. Harvard University Press. 404 p.
Nogueira-Neto, P. 1997. Vida e criação de abelhas indígenas sem ferrão. Nogueirapis, São Paulo. 446p.
Nogueira-Neto, P. 1999. Stressful situation and their consequences on sex determination and on the behavior of stingless bees (Meliponinae, Apidae, Hymenoptera). Revista de Ecologia, 1 (1): 65-68.
Sakagami S F, Laroca S. 1963. Additional observations on the habits of the cleptobiotic stingless bees the genus Lestrimelitta Friese. Journal of Faculty Science of Hokkaido University, Seri I. 15:319-339.
Sakagami S F, Roubik D W, Zucchi R. 1993. Ethology of the robber stingless bee, Lestrimelitta limao (Hymenoptera: Apidae). Sociobiology, 21(2): 237-277.
Wittmann, D. 1985. Aerial defense of the nest by workers of the stingless bee Trigona (Tetragonisca) angustula (Latreille) (Hymenoptera, Apidae). Behaviour Ecology and Sociobiolgy 16:111-114.
Wittmann, D.; Radtke, R.; Zeil, J.; Lubke, G. and Francke, W. 1990. Robber bees (Lestrimelitta limao) and their host chemical and visual cues in nest defense by Trigona (Tetragonisca) angustula (Apidae: Meliponinae). Journal of Chemical Ecology 16(2):631-641.