Artigo
TRANSFERÊNCIA DE COLÔNIAS DE MELIPONÍNEOS PARA COLMEIAS MODELO "UBERLÂNDIA" (Hymenoptera, Apidae, Meliponinae)
AIDAR, Davi S.¹ & KERR, Warwick E.²
1 - Universidade Federal do Amazonas/UFAM, e-mail: aidar@ufam.edu.br
2 - Instituto Nacional de Pesquisas da Amazônia/INPA. Av. André Araújo, 1756, Petrópolis, Cx.P. 478, Cep. 69083-000, Manaus, AM
ABSTRACT
Transfer of meliponíneos colonies into "Uberlândia" beehives (Hymenoptera, Apidae, Meliponinae)
The paucity of pollinating agents complicates the agricultural yield and the maintenance of the bio-diversity on Earth. Meliponinae (Hymenoptera, Apidae, Meliponinae) pollinate up to 90% of all tropical plant species in the tropics and are being extinguished due to the changes on the tropical ecosystems, brought about by the increase on the human population. The transportation of these bees enclosed in their natural substract, e. g., hollow tree trunks, is more difficult than when done in man made beehives. The volume of these beehives still hasn't been well defined for each bee species. Further studies are needed in order to assess the efficiency of each new man made beehive model according to its adaptability to every bee species colony. We surveyed 25 colonies of Melipona compressipes fasciculata Smith (species #4); 3 colonies of Melipona quadrifasciata anthidioides Lep. (species #2); 9 colonies of Melipona marginata Lep. (species #3) and 6 colonies of Scaptotrigona postica Lep. (species #1) relative to the "Uberlândia" man made beehives and the adopted management. The colonies were transferred from the original tree trunks into the "Uberlândia" beehives and ranked according to their adaptability. Grades from 1.0 to 10.0 where assigned to each colony: Grade one to the ones ranked before the transfer and grade 2 to the ones ranked 3 days after the transfer. The average grade fluctuations of species #1, #2, #3 and #4 were -0.33; -0.37; -1.51 and 0.8, respectively. Species #3 ranked according to grade 2 was the one which developed the least, showing the volume of "Uberlândia" beehives was too large for this species, the same happened to species #2. Species #1 and #4 had a better adaptation following the transfer into the "Uberlândia" beehives, pointing out these beehives provided better breeding conditions for these species.
Key Words: man made beehives; Melipona; transfer and tree trunks
1. INTRODUÇÃO
A escassez de agentes polinizadores dificulta a produção agrícola e a manutenção da biodiversidade no Planeta (HOYT, 1992). Os meliponíneos polinizam até 90% das espécies vegetais nos trópicos (KERR et al., 1994) e estão sendo extintos devido às modificações nos ecossistemas tropicais, causadas pelo aumento da população humana, que vem modificando o ambiente e prejudicando a sobrevivência de outras espécies.
O pesquisador que consegue manter 50 matrizes de Melipona, da mesma espécie, consegue superar as dificuldades para obter material biológico para os experimentos. O custo de manutenção do meliponário é o mesmo para um biotério de animais de laboratório. Instalações sofisticadas, suporte técnico e pessoal especializado são fundamentais para a manutenção do biotério. A falta de floradas, os desmatamentos, a prática da monocultura na agricultura e a ação predatória dos tronqueiros ou meleiros, que cortam ilegalmente árvores centenárias para colher o mel e as abelhas, vendendo-os como alternativa de renda, agravam a situação. Laboratórios de pesquisas científicas, nacionais e estrangeiros, são compradores destas abelhas, sendo que deveriam criá-las e produzir colônias para os experimentos sem a necessidade de retirá-las da natureza sempre que precisam.
As pesquisas sobre manejo de meliponíneos para a manutenção da população geneticamente ativa com, no mínimo, 6 alelos xo (44 colônias) evitando a consangüinidade (LAIDLAW et al., 1956; YOKOYAMA & NEI, 1979; KERR & VENKOVSKI, 1982), a multiplicação e alimentação artificiais (AIDAR et al., 1995; AIDAR & CAMPOS, 1994) e os novos modelos de colmeias racionais (KERR, 1995; NOGUEIRA-NETO, 1970 e 1993), que permitem melhor desenvolvimento das colônias, têm possibilitado a sobrevivência das abelhas nas universidades, ou em meliponários particulares, facilitando os estudos e o entendimento da biologia e conservação das espécies.
A avaliação numérica de colônias de meliponíneos (KERR, 1987; AIDAR, 1995) auxilia a padronização dos dados de experimentos de diferentes laboratórios, facilitando o acompanhamento dos efeitos causados por diferentes técnicas de manejo, transferência, transporte e divisões de colônias.
O transporte de meliponíneos em seus substratos naturais (troncos de árvores ou cortiços) sempre é mais difícil do que o transporte em colmeias racionais com tamanhos padronizados e de menor peso. A transferência das colônias do tronco para colmeias racionais, dias antes de se realizar o transporte, permite o acondicionamento de um número maior de colmeias num mesmo veículo, sem provocar escape de abelhas ou mesmo morte por falta de arejamento, diminuindo os custos, o tempo do transporte e morte de colônias.
O volume das colmeias racionais para cada espécie de meliponíneo ainda não está bem definido, porém alguns dados podem proporcionar melhores rendimentos às colônias: para Melipona scutellaris Lep. e Melipona compressipes fasciculata Smith, KERR (1987) sugere 27 litros; para Melipona beecheii, QUEZADA-EUAN & GONZALES-ACERETO (1994); para Melipona quadrifasciata Lep., Melipona favosa orbigini Lep. e Tetragonisca angustula, AIDAR (1996) sujere 14, 10 e 8 litros, respectivamente.
Existem muitos modelos de colmeias racionais, desde os mais primitivos até os mais modernos (NOGUEIRA-NETO, 1970) que proporcionam um melhor aproveitamento dos subprodutos das abelhas. Vários modelos de colmeias para meliponíneos são citados em literatura, porém necessitam de estudos com repetições suficientes que permitam análise estatística para a comprovação da eficiência de cada modelo com relação ao desempenho das espécies de abelhas.
Neste trabalho procurou-se avaliar as colônias de diferentes espécies com relação à colmeia modelo "Uberlândia" (KERR et al., 1996) e ao manejo aqui adotado: Melipona compressipes fasciculata Smith (uruçu); Melipona quadrifasciata anthidioides Lep. (mandaçaia); Melipona marginata Lep. (manduri) e Scaptotrigona postica Lep. (mandaguari).
2. MATERIAL E MÉTODOS
Os experimentos foram realizados em Prudentópolis, PR, Mearim e Arari, MA. Melipona comprescipes fasciculata (tiúba), Melipona quadrifasciata anthidioides (mandaçaia), Melipona marginata (manduri) e Scaptotrigona postica (mandaguari) foram as 4 espécies estudadas. Vinte e cinco, 4, 9 e 3 colônias foram o número de repetições para cada espécie, respectivamente. As colônias foram transferidas do tronco para colmeias racionais modelo "Uberlândia".
No sentido de avaliar o desempenho das colônias, foram atribuídas notas de 1,0 a 10,0 antes da transferência (Nota 1) e 3 dias após a transferência (Nota 2). No caso das Melipona compressipes houve diferenciação na metodologia com relação ao período entre revisões e o número de revisões. Foram realizadas 4 inferências de notas para esta espécie: Nota1, imediatamente após a transferência; Nota 2, 10 dias após; Nota 3, 33 dias após e Nota 4, 46 dias após a inferência da Nota1.

Figura 1. Vista superior de uma colmeia racional preparada com cera de abelhas
Apis mellifera para receber a colônia de Melipona, mostrando o invólucro (i)
e o tubo de entrada (t).
As colmeias foram preparadas antes da transferência com a construção de um invólucro de crias e um tubo de entrada confeccionados com lâminas de cera alveolada de abelhas africanizadas (Figura 1). Em seguida, a entrada da colônia foi transferida para a colmeia racional. Como a entrada natural da colmeia de meliponas é feita de geoprópolis e sua remoção é muito difícil sem que seja danificada, esta foi macerada e umidificada de forma a obter-se uma pasta. Esta pasta foi colocada no orifício da colmeia de forma a ficar totalmente tapado e, com auxílio de um lápis, foi feito novo orifício. Apenas um pedaço da entrada original poderá ser transferida, sendo suficiente para que as campeiras localizem a entrada da nova moradia; porém, este pedaço deve ser fixo com fita adesiva ou cera da própria colônia, o que é preferível. No caso de abelhas do Gênero Trigona, a entrada da colmeia é feita de cera e pode ser transferida com certa facilidade para a colmeia racional, sem que se desmanche.
O fundo da colmeia foi fixo com fita adesiva e a tampa também foi vedada pelo mesmo sistema, mas somente após a transferência ter sido completada.
Preparada a colmeia racional, o tronco foi serrado longitudinalmente de ambos os lados, com auxílio de moto-serra, sem que o oco onde estava o ninho fosse atingido pela corrente da moto-serra. Com auxílio de cunhas e marreta, o tronco foi partido ao meio, longitudinalmente e foram transferidos para colmeia os favos de crias e os potes de pólen e mel. Quando necessário, foram colocados potes artificiais confeccionados com cera de abelhas africanizadas de acordo com AIDAR (1995 e 1996) e adicionado suplemento com xarope-A (AIDAR, 1996).
As abelhas jovens que não podem voar e durante o processo de transferência caem no chão ou ficam andando pelo tronco, foram recolhidas com auxílio de um aspirador de insetos e colocadas dentro da colmeia.
A rainha fisogástrica foi colocada em placa de Petri ou gaiola Doolitle até o processo de transferência ter sido completado. A rainha foi colocada sobre os favos de crias, e a colmeia fechada, a tampa vedada com fita adesiva e a colmeia foi colocada no lugar do tronco de forma que a entrada coincidisse com a entrada original (entrada do tronco). Uma cobertura com telha de cimento amianto foi colocada sobre a colmeia para evitar raios solares diretamente às suas paredes e umidade decorrente do sereno e das chuvas. Os raios solares diretamente na colmeia promove superaquecimento e derretimento de cera dentro da colmeia.
Nas colmeias onde a infestação por forídeos foi acentuada, foram colocadas armadilhas internamente e externamente para reduzir a infestação. Estas armadilhas não resolvem o problema, mas podem amenizá-lo se bem manejadas.
3. RESULTADOS E DISCUSSÃO
Entre a Nota 1 e a Nota 2 avalia-se o efeito da transferência do tronco para a colmeia racional, conforme metodologia empregada (a ser descrita). Após a Nota 2, avalia-se o desempenho das colônias nas colmeias racionais sem o efeito do stress da transferência, ou seja, avalia-se a adaptação das abelhas com relação à colmeia utilizada no experimento.
Para as mandaguaris (Espécie 1), mandaçaias (Espécie 2) e manduris (Espécie 3), as notas antes da transferência (N1) e após a transferência (N2), estão apresentadas na Tabela1.
| ESPÉCIE 1 | ESPÉCIE 2 | ESPÉCIE 3 | ||||
| COLÔNIA | Mandaguari | Mandaçaia | Manduri | |||
| N1 | N2 | N1 | N2 | N1 | N2 | |
| 1 | 6,5 | 6,0 | 6,0 | 5,0 | 7,0 | 3,0 |
| 2 | 8,0 | 8,0 | 4,0 | 3,8 | 7,0 | 4,5 |
| 3 | 8,0 | 7,5 | 4,0 | 4,0 | 7,0 | 5,0 |
| 4 | 7,0 | 4,0 | 4,0 | 2,8 | ||
| 5 | 5,0 | 3,2 | ||||
| 6 | 6,0 | 4,0 | ||||
| 7 | 4,0 | 3,0 | ||||
| 8 | 6,0 | 4,8 | ||||
| 9 | 4,0 | 2,5 | ||||
| N2-N1 | ||||||
| X1 | -0,5 | -0,1 | -0,4 | |||
| X2 | 0,0 | -0,2 | -2,5 | |||
| X3 | -0,5 | 0,0 | -2,0 | |||
| X4 | -1,2 | |||||
| X5 | -1,8 | |||||
| X6 | -2,0 | |||||
| X7 | -1,0 | |||||
| X8 | -1,2 | |||||
| X9 | -1,5 | |||||
| ÓN | -0,1 | -1,5 | -13,6 | |||
| x | (n=3) -0,333 | (n=4) -0,375 | (n=9) -1,510 | |||
| S2x | 0,084 | 0,189 | 0,404 | |||
| Sx | 0,290 | 0,455 | 0,635 | |||
| Sx | 0,170 | 0,220 | 0,210 | |||
Onde: εn = soma da diferença da nota; x = média; s2x = variância de x; Sx = desvio padrão de x; sx = desvio padrão da média.
Nesta segunda avaliação os fatores que mais influenciam o desempenho das colônias são: efeito do genótipo (linhagem) e efeito da rainha (idade), neste caso, o efeito do estresse da transferência está excluído pois passaram-se 63 dias e as abelhas já se reestruturaram dentro da nova moradia.
Na Figura 2 está representado o Diagrama de Pontos das espécies 1, 2 e 3, mostrando a distribuição das variações das notas.
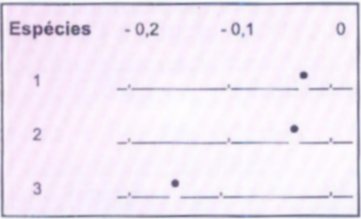
Figura 2. Diagrama de pontos ilustrando a distribuição
média das variações das notas das espécies estudadas.
Verifica-se que as espécies 1 e 2 têm variação entre -1 e 0, enquanto que a espécie 3 varia entre -2,5 e -1,0, com exceção da colônia 1 com diferença de -0,4 pontos da nota, considerando a proximidade da média das espécies 1 e 2 e a pouca repetição, pôde-se construir intervalos de confiança agrupando estas duas espécies:
Intervalos de confiança para a média da diferença entre as notas: I.C.(90%) = x±(ta/2.sx)
Esp.1 -0,829; +0,163
Esp.2 -0,892; +0,142
Esp.(1 e 2) -0,613; -0,101; onde n=7; x=-0,357 e sx=0,132
Esp.3 -1,900; -1,120
As médias da variação das notas das espécies 1, 2 e 3 foram -0,33; -0,37 e -1,51, respectivamente. As manduris apresentaram N2 menor, o que sugere volume excessivo das colmeias para esta espécie, já que as manduris apresentam colônias com menor população entre as espécies aqui estudadas. Quando o volume da colmeia é maior do que o ótimo para a espécie em questão, as colônias apresentam crescimento lento com vulnerabilidade a ataques de predadores, principalmente forídeos (Pseudohipocera sp) e às variações da temperatura ambiente. A espécies 1, após 3 dias adaptaram-se bem ao modelo de colmeia utilizada. Esta espécie é caracterizada por grande número de indivíduos por colônia e maior quantidade de favos de crias, quando comparadas às outras 3 espécies, proporcionando ocupação rápida dos espaços internos da colmeia e reestruturação precoce dos danos causados pela manipulação durante os trabalhos de transferência da colônia. Para a espécie 2, sugere-se um volume de no máximo 15 litros (AIDAR, 1995) e não 27 litros das colmeias modelo "Uberlândia" aqui utilizadas. Mesmo assim, foi a espécie 2 recuperou-se relativamente bem quando comparada às outras espécies.
| COLÔNIA | N1(29/7/81) | N2(8/8/81) | N3(31/8/81) | N4(13/9/81) | n Obs. |
| K60 | 2,0 | 5,5 | 5,0 | 5,0 | 1 |
| K43 | 8,0 | 6,5 | 8,0 | 7,0 | 2 |
| K58 | 6,0 | 5,5 | 5,0 | 5,0 | 3 |
| K59 | 6,0 | 6,0 | 7,0 | 8,0 | 4 |
| K45 | 3,0 | 0,5 (s/rainha) | 0,0 | 0,0 | 5 morre |
| K56 | 4,0 | 4,5 | 4,0 | 4,0 | 6 |
| K48 | 7,0 | 9,0 | 10,0 | 9,0 | 7 |
| K42 | 6,0 | 6,0 | 9,0 | 7,0 | 8 |
| K50 | 4,0 | 5,0 | 8,0 | 6,0 | 9 |
| K54 | 9,0 | 9,0 | 9,5 | 8,0 | 10 |
| K51 | 6,0 | 7,5 | 6,0 | 7,0 | 11 |
| K44 | 6,0 | 6,0 | 9,0 | 9,0 | 12 |
| K46 | 6,0 | 7,5 | 8,0 | 8,0 | 13 |
| K47 | 5,0 | 1,0 (s/rainha) | 6,0 | 6,0 | 14 |
| K52 | 7,0 | 10,0 | 10,0 | 8,5 | 15 |
| K49 | 6,0 | 10,0 | 8,0 | 6,0 | 16 |
| K55 | 4,0 | 10,0 | 8,5 | 6,0 | 17 |
| K41 | 4,0 | 5,5 | 4,0 | 4,0 | 18 |
| W66 | 5,0 | 8,0 | 9,5 | 9,0 | 19 |
| W65 | 7,0 | 6,5 | 6,5 | 4,0 | 20 |
| W64 | 6,0 | 6,5 | 8,5 | 7,0 | 21 |
| W67 | 5,0 | 6,0 | 8,5 | 7,5 | 22 |
| K53 | 3,0 | 4,0 | 5,0 | 4,5 | 23 |
| W61 | 6,0 | 4,0 | 4,0 | 5,0 | 24 |
| W62 | 2,0 | 3,0 | 3,0 | 4,0 | 25 |
No caso da espécie 4, as notas N1, N2, N3, e N4 estão apresentadas na Tabela 2, bem como a identificação das colônias e outras observações.
A primeira análise estatística realizada foi a correlação N1 e N2:
1) Correlação N1 e N2: A = 1,990 (ponto onde a reta corta o eixo Y)
B = 0,777 (inclinação da reta)
R = 0,540
n = 25; gl=23
x = 5,32 (N1)
y = 6,12 (N2)
x = 1,75 (erro da média)
y = 2,51 (erro da média)
y - x = 0,80 (diferênça entre as médias)
2) Correlação N1 e N4: A = 2,370 (ponto onde a reta corta o eixo Y)
B = 0,720 (inclinação da reta)
R = 0,59
n = 25; gl = 23
x = 5,32 (N1)
y = 6,18 (N2)
x = 1,75 (erro da média)
y = 2,12 (erro da média)
y - x = 0,86 (diferênça entre as médias)
Pela tabela da distribuição t, gl = 23; é significativo a 10%.
Pode ser que em 2 ou 3 dias a espécie 4 tenha tido decadência igual às outras 3 espécies estudadas. Devemos observar que a N2 é 10 dias após, enquanto que para as espécies 1, 2 e 3, foi de 2 dias apenas. Mesmo assim, a média de variação das diferenças das notas de cada revisão foram positivas: N2 - N1 = 0,8; N3 - N1 = 1,48 e N4 - N1 = 0,86. O que mostra que as colônias apresentam crescimento, com exceção dos casos críticos da K45 e K47 que ficaram sem rainha e tiveram a nota em queda. A K45 morreu por não ter substituído a rainha fisogástrica, que fora morta, e a K47 recuperou-se após a substituição da rainha.
Estudando os intervalos de confiança para N2 - N1, N3 - N1 e N4 - N1, obteve-se os seguintes resultados para N2 - N1: D = 0,80; S = 2,20; S2 = 4,84; SD = 0,44 e I.C.90%(D) = 0,89±0,40x1,72 = (0,29; 1,67), sendo que, neste caso, excluiu-se as colmeias K45 e K47, que ficaram sem rainhas, para não interferirem na média geral da maioria das colônias (n = 23), já que, estas colmeias juntas representam apenas 8% do total de repetições.
Para N3 - N1: D = 1,48; S = 1,98; S2 = 3,94; SD = 0,40 e I.C.90%(D) = 1,48±0,40x1,71 = (1,08; 2,16).
Para N4 - N1: D = 0,86; S = 1,78; S2 = 3,18; SD = 0,35 e I.C.90%(D) = 0,86±0,35x1,71 = (0,51; 1,46).
Onde D = média da diferença das notas; S = desvio padrão; S2 = variância; SD = desvio padrão da média e I.C.90%(D) = D±S x t. Sendo t = 1,72; 1,71 e 1,71, respectivamente para os três intervalos calculados.
Percebe-se que o intervalo se estreita com o tempo, isto é, a variação das diferenças das notas (d) diminui com o tempo: d (N2 - N1) = 1,38; d (N3 - N1) = 1,08 e d (N4 - N1) = 0,95. Isto pode ser explicado pela homogeneidade do ambiente: mesmo manejo; mesmas colmeias e mesmo local). O que, comumente não ocorre quando as colônias estão na natureza, onde cada oco de árvore apresenta características próprias: tipo de árvore; espessura da parede do oco; localização da árvore (sol, sombra, umidade, vento, formigas, etc.); entre outros. Deve ser lembrado que F = G + A; onde F = fenótipo; G = genótipo e A = ambiente. No caso, padronizou-se as colmeias e o manejo, ou seja, o ambiente (A), proporcionando às colônias condições de melhor expressão fenotípica (F) de acordo com os seus genótipos (G) e seqüente desempenho quanto à produtividade com relação aos elementos que compõem uma colônia de meliponíneo (AIDAR, 1996): número de campeiras; invólucro do ninho; postura da rainha fisogástrica; potes de alimento; entrada da colmeia; resina coletada; favos de crias; organização geral das abelhas dentro da colmeia e ganho de peso da colmeia.
4. CONCLUSÕES
As uruçus não apresentaram variação negativa das notas após a transferência para colmeias racionais, mostrando que a colmeia ofereceu boa adaptabilidade para esta espécie.
As mandaçaias requerem volume menor de colmeia para um bom desempenho das colônias. Os forídeos não ofereceram grandes transtornos para esta espécie, no 3o dia já não haviam os dípteros dentro das colmeias de mandaçaias.
As manduris apresentaram pior desempenho, supondo que o volume da caixa foi excessivo para estas abelhas, que apresentam população menor e mais sensibilidade às variações ambientais. Já as mandaguaris, por apresentarem população maior e mais favos de crias com relação às outras espécies estudadas, apresentaram boa adaptabilidade às colmeias mesmo apresentando vulnerabilidade aos forídeos. Isto ocorre porque esta espécie apresenta pólen com odor mais forte e adocicado, o que atrai mais forídeos que as outras colônias.
Os forídeos foram freqüentes mas bem controlados com o manejo adotado e com o auxílio de armadilhas. A infestação maior ocorreu no 2o dia após a transferência. O local onde os forídeos foram mais coletados foi quando as armadilhas foram dispostas externamente à entrada da colmeia. Internamente às colmeias, as armadilhas não surtiram bons resultados.
Agradecimentos
Agradecemos à FAPEMIG (Fundação de Amparo à Pesquisa de Minas Gerais, ao CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) e à Fundação Banco do Brasil pelos auxílios financeiros. À Dra. Tirza Aidar da UNICAMP (Universidade de Campinas) pela revisão nas análises estatísticas.
6. BIBLIOGRAFIA
AIDAR, Davi S. 1996. A mandaçaia: Biologia de abelhas, manejo e multiplicação artificial de colônias de Melipona quadrifasciata Lep. (Hymenoptera, Apidae, Meliponinae). Série Monografias 4, Bras. J. Genetics, 103p.
AIDAR, Davi S. & CAMPOS, Lucio A.O. 1994. Resposta de meliponíneos à alimentação artificial: Melipona quadrifasciata Lep. (HYMENOPTERA, APIDAE,). Ann. Enc. Etologia, 12:105-106.
AIDAR, Davi S.; CAMPOS, Lucio A.O.; POMPOLO, Silvia G.; MESSAGE, Dejair; AIDAR, Tirza. 1995. Influência Ambiental na multiplicação artificial de meliponíneos: Melipona quadrifasciata Lep. (HYMENOPTERA, APIDAE). In: Ann. Congr. de Entomologia, 15, Caxambú, MG.
HOYT, Erich. (1992). Conservação dos parentes silvestres das plantas cultivadas. Trad.Lídio Coradfin. ADDISON-Wesley IBEROAMERICANA, Wilmington, Delaware, III/S.A. 52pp.
KERR, Warwick E.; CARVALHO, Gislene A. e NASCIMENTO, Vânia A. 1996. A abelha uruçu: Biologia, manejo e conservação. Fundação Acangau, Belo Horizonte, MG, Brasil. 222p.
KERR, Warwick E. 1987. Biologia, manejo e genética de Melipona compressipes fasciculata Smith (HYMENOPTERA, APIDAE). Univ. Fed. Maranhão, São Luiz, MA. 141p. (Tese Titular).
KERR, Warwick E.; NASCIMENTO, Vânia A. e CARVALHO, Gislene A. 1994. Há salvação para os meliponíneos? In: Anais Enc. sobre Abelhas, 1:60-65, Ribeirão Preto, SP.
KERR, Warwick E. & VENCOVSKY, Roland.1982. Melhoramento genético em abelhas I: Efeito do número de colônias sobre o melhoramento. Rev. Brasil. Genét. 5:279-285.
LAIDLAW, H.H.; GOMES; F.P. e KERR, W.E. 1956. Estimation of the number of lethal alleles in a panmitic population of Apis mellifera L. Genetics 44:179-188.
NOGUEIRA-NETO, Paulo. 1970. A criação de abelhas indígenas sem ferrão (Meliponinae). 2a ed., Tecnapis, São Paulo, SP. 365p.
NOGUEIRA-NETO, Paulo. 1993. Novas técnicas para criar meliponíneos (HYMENOPTERA, APIDAE, MELIPONINAE). Publ. Tecnapis sobre Ecologia e Etologia (Ecoetologia), 5, 15p.
QUEZADA-EDUÁN, José Javier G.; GONZALEZ-ACERETO, Jorge. 1994. A preliminary study on the development of colonies of Melipona beecheii in traditional and rational hives. Journal of Apic. Research 33(3):167-170.
YOKOYAMA, S. & NEI, M. 1979. Population dynamics of sex-determinning alleles in honey bees and self-incompatibility alleles in plants. Genetics 91:609-626.